
Gametofiton/sporofiton nemzedékek a növényvilágban
A szárazföldi növényvilágban általános egyedfejlődés-típus a nemzedékváltakozás, vagy más néven a kétszakaszos egyedfejlődés. Egy növény életében mindig megjelenik időben egymást követve annak kétféle kromoszóma számú formája. Az egyedfejlődésében mindig szabályszerűen megjelenik a haploid spórából kifejlődő, feles „n”-kromoszóma számú sejtekből álló, ivarszerveket és ivarsejteket létrehozó ivaros nemzedék a gametofiton. E nemzedék haploid ivarsejtjeinek egyesülésével létrejön egy diploid zigóta, amelyből mitotikus osztódásokkal egy 2n kromoszómás szervezet fejlődik ki. Ezen jönnek létre a diploid spóraanyasejtek, amelyek meiozissal haploid spórákat hoznak létre. Innen származik e diploid nemzedék neve: sporofiton..
Megjegyzendő, hogy a „nemzedék” szó használata nem igazán szerencsés, hisz ebben a vonatkozásban a fogalom tartalma nem genetikai, hanem egy adott egyed fejlődésének két egymást követő, egymást feltételező, egymásra épülő szakaszát jelenti. A két szakasz élesen elhatárolódik a nemzedékek sejtjeinek eltérő kromoszóma garniturája miatt/haploid gametofiton és diploid sporofiton/. Azonban a „nemzedék” szó, a nemzedékváltakozás fogalom annyira rögzült a magyar növénytani szakzsargonban, hogy általánosan elfogadható helyettesítése valami más kifejezéssel reménytelennek tűnik.
Közismert, hogy a természetben kétféle ivarsejt létezik. A rendszerint kisebb, mozgékony hím ivarsejt és a nagyobb mozdulatlan női ivarsejt/petesejt/. A kétféle gaméta és az őket létre hozó két különböző ivarszerv fejlődhet egy gametofiton, vagy alakilag is elválhatnak egymástól. A szárazföldi növényeknél már a harasztoknál is megfigyelhetünk olyan eseteket, amikor külön fejlődik egymástól a kétféle ivaros nemzedék. A kisebb mikrospórából kifejlődő mikrogametofiton hozza létre a a hím ivarszerveket/antheridium/ és ebben a hímivarsejteket. A nagyobb makrospórákból fejlődő makrogametofiton a női ivarszerv/archegonium/ és a petesejt kifejlődésének helyszíne. A kétféle gametofiton elkülönült fejlődése a virágos, magvas növényeknél/nyitva-,és zárvaternők/ általános.
A szárazföldi növényeknél a gametofiton és a sporofiton aránya változó Általános tendencia, hogy a törzsfejlődés során a gemetofiton egyre kisebb lett, redukálódott. A moháknál még a zöld leveles, vagy teleptestű közismert növényke a gametofiton.  A mohákon később megjelenő barna nyél és a tetején kifejlődő spóratermelő kupakos tok az ivartalan szaporítósejteket/spórákat/ létrehozó sporofiton nemzedék. Ebben a növénycsoportban a sporofiton lényegében a gametofiton nemzedéken élősködik.
A mohákon később megjelenő barna nyél és a tetején kifejlődő spóratermelő kupakos tok az ivartalan szaporítósejteket/spórákat/ létrehozó sporofiton nemzedék. Ebben a növénycsoportban a sporofiton lényegében a gametofiton nemzedéken élősködik.
 A harasztoknál a gametofiton már jóval kisebb, szabad szemmel éppen csak látható. Az ún. előtelep/protallium/ egy csoporttól függő külsejű lemezes képződmény. A páfrányoknál például szív alakú /fotoszintézisre is képes lehet/, melyen az ivarszervek és bennük az ivarsejtek fejlődnek.
A harasztoknál a gametofiton már jóval kisebb, szabad szemmel éppen csak látható. Az ún. előtelep/protallium/ egy csoporttól függő külsejű lemezes képződmény. A páfrányoknál például szív alakú /fotoszintézisre is képes lehet/, melyen az ivarszervek és bennük az ivarsejtek fejlődnek.

Páfrány előtelep-gametofiton
Az ivarsejtek egyesülése/megtermékenyítés/ általában a víz közvetítésével és kemotaktikus ingerek/pl almasav/ irányításával az előtelepen történik. A sporofiton nemzedék első sejtje/a diploid zigóta/ is az előtelepen jelenik meg. A belőle fejlődő sporofiton kialakulása itt a protalliumon indul meg. Továbbiakban az elpusztuló gametofiton nemzedéken kezdik el fejlődésüket a valódi hajtásos, de virágtalan harasztnövények/korpafüvek, zsurlók,páfrányok/ sporofiton alakjai, amelyek leveleik különböző részein, vagy elkülönült spóratermelő szerveikben később majd spórákat hoznak létre.

Páfrány sporofiton sporangiumokkal és szóruszokkal
 A nyitva-, és zárvatermők ivaros nemzedéke tovább egyszerűsödik. Míg a korpafüvek és a harasztok előtelepe (protalliuma) még teljesen önálló, lemezes szerveződésű, autotrof, – fotoszintézisre képes – ivarszerveket létrehozó forma, addig a virágos növényekben az ivaros nemzedék teljesen elveszíti önállóságát. A virágos, magvas növényeknél (Spermatophyta) mindkét gametofiton a virágban a diploid sporofiton védelmében fejlődik. Attól kapja a tápanyagait is, önálló autotrof anyagcserére képtelen. A növényvilág evolúciója során a női ivarsejteket termelő ún. makrogametofiton mindig fejlettebb marad, mint a hím ivarsejteket létrehozó mikrogametofiton.
A nyitva-, és zárvatermők ivaros nemzedéke tovább egyszerűsödik. Míg a korpafüvek és a harasztok előtelepe (protalliuma) még teljesen önálló, lemezes szerveződésű, autotrof, – fotoszintézisre képes – ivarszerveket létrehozó forma, addig a virágos növényekben az ivaros nemzedék teljesen elveszíti önállóságát. A virágos, magvas növényeknél (Spermatophyta) mindkét gametofiton a virágban a diploid sporofiton védelmében fejlődik. Attól kapja a tápanyagait is, önálló autotrof anyagcserére képtelen. A növényvilág evolúciója során a női ivarsejteket termelő ún. makrogametofiton mindig fejlettebb marad, mint a hím ivarsejteket létrehozó mikrogametofiton.
A nyitvatermők közös sajátossága, hogy ivaros nemzedékeik újszerűen már módosult leveleken fejlődnek. A női ivaros nemzedék/makrogametofiton/ a termős toboz módosult levelén a makrosporofillumon/termőlevél/ fejlődik. Azonban ez csak részleges védelmet biztosít a női gametofiton számára, mert a fenyőféléknél a termőlevelek a zárvatermőkkel ellentétben nem forrnak össze egy zárt magházzá. A női ivaros nemzedék az ivarleveleken egy új szervben az un magkezdeményekben fejlődik, melyek többé kevésbé szabadon állnak a termős tobozok termőlevelein.
A női ivaros nemzedék/makrogametofiton/ a termős toboz módosult levelén a makrosporofillumon/termőlevél/ fejlődik. Azonban ez csak részleges védelmet biztosít a női gametofiton számára, mert a fenyőféléknél a termőlevelek a zárvatermőkkel ellentétben nem forrnak össze egy zárt magházzá. A női ivaros nemzedék az ivarleveleken egy új szervben az un magkezdeményekben fejlődik, melyek többé kevésbé szabadon állnak a termős tobozok termőlevelein.

Fenyő magkezdemények
A női makrogametofiton a fenyőknél alapvetően eltér a zárvatermők hasonló haploid nemzedéktől. A toboz termőlevelek tövében a magkezdemények közepén egy makrospórából alakul ki a mikroszkópikus méretű előtelep, amely több ezer sejtből áll. Lényeges különbség a két növénycsoport között, hogy a nyitvatermők makroprotalliumában még kialakul egy archegónium (női ivarszerv), ami a zárvatermőknél teljesen hiányzik. Ebben alakul ki a petesejt, amelyet a pollenből fejlődő hím ivaros nemzedék hím ivarsejtje/spermatozoid/ termékenyít meg.
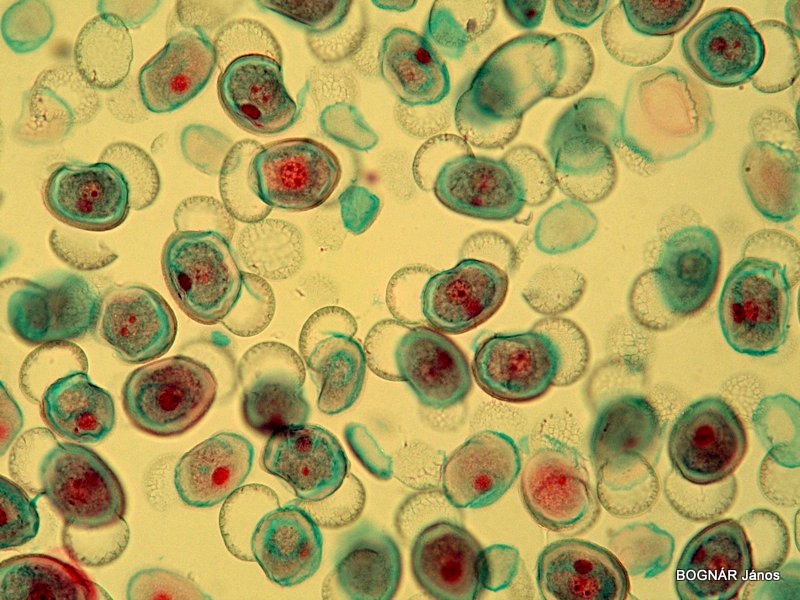
Fenyő pollenszemek
A hím mikrogametofiton fejlődése is bonyolultabb, mint a zárvatermőké, még egy sejtosztódási folyamat és néhány plusz sejt még beiktatódik. Megjelenik a két hímivarsejt, de általában nincs kettős megtermékenyítés.
A zárvatermők evolúciós újítása a virágban a termőlevelekből kialakuló zárt magház, melynek belsejében a magkezdeményekben fejlődik a női ivaros nemzedék. A teljes női ivaros nemzedék a magkezdeményben helyet foglaló embriózsákban elfér. A női gametofitont alkotó sejtek száma a nyitvatermőkhöz viszonyítva itt még tovább csökken.  A leggyakrabban emlegetett ősi un normál „Polygonum-típusú” női gametofinnál az embriózsák lényegében egy 8 haploid sejtből fejlődő különös „szupersejt”,
A leggyakrabban emlegetett ősi un normál „Polygonum-típusú” női gametofinnál az embriózsák lényegében egy 8 haploid sejtből fejlődő különös „szupersejt”,  amely a fejlődési folyamat végén már csak 7 sejtmagot tartalmaz/un szekunder embriózsák/: egy centrális fúzionált diploid központi vegetatív magot, és hat haploid sejtmagot: az egyik oldalon csoportosulva egy petesejtet és annak két kísérő sejtjét, az ellentétes oldalon pedig még három ellenlábas sejtet. Az osztódásokat követően és a végleges poziciók kialakulása után megalakul a magokat elkülönítő sejtfal is a diploid központi sejtet kivéve, ahol kifejlett állapotban sincs határoló felület a központi magpár körül. A női előtelep lényegében az ellenlábas sejtek alkotta trió, az ivarsejteket a petesejt és a”biztonsági tartalék”-nak megfelelő két kísérősejt reprezentálja.
amely a fejlődési folyamat végén már csak 7 sejtmagot tartalmaz/un szekunder embriózsák/: egy centrális fúzionált diploid központi vegetatív magot, és hat haploid sejtmagot: az egyik oldalon csoportosulva egy petesejtet és annak két kísérő sejtjét, az ellentétes oldalon pedig még három ellenlábas sejtet. Az osztódásokat követően és a végleges poziciók kialakulása után megalakul a magokat elkülönítő sejtfal is a diploid központi sejtet kivéve, ahol kifejlett állapotban sincs határoló felület a központi magpár körül. A női előtelep lényegében az ellenlábas sejtek alkotta trió, az ivarsejteket a petesejt és a”biztonsági tartalék”-nak megfelelő két kísérősejt reprezentálja.
Ettől az általánosan elterjedt típustól eltérő változatokat is találunk. Az „Oenothera”- típus mindössze 4 sejtes, hiányzik a diploid központi sejt és a két kísérősejt. A másik végletet megjelenitő „Panea” típusnál a kész szekunder embriózsák 16 sejtből áll. A variációk zöme azonban a normál ősi típushoz hasonlóan nyolc sejtes. A különbségek oka az osztódások kezdetén a kiinduló spórák eltérő számában/mono-, bi-,és teraspórás eredet/ és a további fejlődési utakban mutatkozó eltérésekben keresendő.
A hím ivaros nemzedék a zárvatermőkben szinte a lehetséges végletekig egyszerűsödik. A redukciós osztódással létrejött mikrospóra/pollenszem/ egyenlőtlenül/inekválisan/ osztódik/un pollenmitózis-1/ és létrejön egy kisebb generatív és egy nagyobb vegetatív magot tartalmazó sejtpáros a kétmagvas pollen. Néhány növénycsoportot kivéve/pl fészkesvirágúak, ernyősök/ a zárvatermők pollenszemei kétsejtes állapotban kerülnek a megporzással a bibére. A pollen itt tömlőt hajt/”csírázik”/. A tömlőben elöl haladó vegetatív sejtet követi a generatív sejt ez utóbbi itt még egyszer osztódik/pollenmitózis-2/. Így alakul ki a két haploid hímivarsejt, amely mai ismereteink szerint nem teljesen egyforma. A sejtmag és a vakuolumok méretében, valamint a proplasztiszok számában különböznek egymástól. Lényegében a vegetatív tömlősejtből és a két hímivarsejtből álló három sejtmagvas pollen a zárvatermők hím ivaros nemzedéke.
A pollentömlő általában a magkezdemény nyílásán/mikropile/ jut az embriózsákba/porogámia/ mindig a kísérősejtek felől. Ritkán egyes növénycsoportokban azonban előfordulhat más lehetőség is/aporogámia/, amikor kerülő úton jut el a petesejt közelébe/diófélék, nyírfák, tökfélék/.
A segítősejtek sejtfalbontó enzimeikkel nyitják fel a pollentömlő falát, ekkor a vegetatív mag elpusztul és két hímivarsejt egyike valószínű amöboid mozgással eléri a petesejtet és fuziójukkal létrejön a diploid nemzedék első sejtje a zigóta, amelyből a továbbikban azembrió fejlődik. A másik hímivarsejt egy passzív folyamattal belökődik az embriózsák közepére és itt egyesül a központi diploid sejtmaggal létrehozva így egy triploid endospermium sejtmagot, amelyből raktározószövet fejlődhet a továbbiakban.
Ez a folyamat a kettős megtermékenyítés, amely zárvatermő növényekre általánosan jellemző és teljes diploid sporofiton fejlődésük kezdőpontja.