
A zsurlók osztályában az Equisetum nemzetség fajainak spórái eredetüket és fő funkciójukat tekintve azonosak a növényvilág többi haploid ivartalan szaporító sejtjével, azonban néhány tulajdonságukban jelentősen különböznek nemcsak más Sphenopsida osztálybeli növény rokontól, hanem az összes többi haraszttól is.
 A zsurlók a külsőre egymáshoz hasonló, de ivarjelleg szempontjából eltérő spóráikat/un homoiospórák/ különálló speciálisan módosult levelek/sporofillumok/ szélein lelógó zsákocskákban/sporangiumok/ hozzák létre redukciós osztódással. A sporofillumok a hajtástengely csúcsán elkülönülve magányos sporofillum füzérben/strobilus/ helyezkednek el.
A zsurlók a külsőre egymáshoz hasonló, de ivarjelleg szempontjából eltérő spóráikat/un homoiospórák/ különálló speciálisan módosult levelek/sporofillumok/ szélein lelógó zsákocskákban/sporangiumok/ hozzák létre redukciós osztódással. A sporofillumok a hajtástengely csúcsán elkülönülve magányos sporofillum füzérben/strobilus/ helyezkednek el.
A gömbölyű spórák kifejlett állapotban etioplasztiszokat és kloroplasztiszokat tartalmaznak, utóbbiak miatt zöldek. Ellenálló, vastag spórafaluk bonyolult felépítésű, három rétegből áll/endo-, mezo- és perispórium/. A legkülső túlnyomórészt cellulóz sejtfalú perispórium a fejlődés kezdetén táplálja a szaporító sejteket, majd a spóraérés végén négy szalagszerű sávra, un hapterákra hasad.
Leclerc du Sablon nevezte el a zsurlók spóráin párosával található szalagokat elateráknak. Szintén ő vizsgálta először a szalagok kémiai felépítését, mikroszkópos szerkezetét. Megállapította, hogy a spórák felületére egy ponton rögzült képződmények elhalt, plazmát nem tartalmazó „üres” sejtekből állnak. Sejtfaluk túlnyomórészt cellulózból épül fel, de a spórához közelebb eső/adaxiális/ belső felületükön a sejtek falai parásodtak, illetve viaszos kutin berakódást tartalmaznak.
Párás térben az elatera külső részében a cellulóz sejtfalak hidratálódnak és megnyúlnak, míg a belső rész a berakódott anyagok miatt nagymérvű vízmegkötésre és kiterjedésre képtelen Az egyenlőtlen megnyúlás következménye a befelé görbülő spirális mozgás.
A cellulózhoz kötött vízmolekulák kiszáradáskor a páratartalom csökkenésével ismét leszakadnak, eldiffundálnak. Emiatt ellenkező mozgás kezdődik és az elaterák legombolyodnak a spóra testéről.
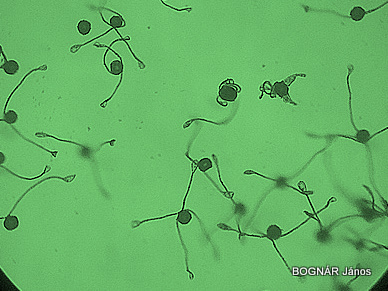
Zsurló homospórák kinyílt elaterákkal
Az elaterák tehát nedves állapotban rácsavarodnak a spórákra, míg száraz állapotban kiterülnek és sajátos összekapcsoló-röpítő készülékként működnek. A spóratartók kiszáradásakor a tárt karokhoz hasonló módon a nyúlványok egymással összeakaszkodnak, így elősegítik a spóratömeg fellazulását, és felületnövelőként a spórák csoportos utazását biztosítják a szelek szárnyain. Egyszerre általában több homoiospóra terjed „társas utazással”, ami azért fontos, mert a külsőre egyforma homospórákból eltérő ivarszerveket kialakító előtelepek fejlődnek.

Spórára tekeredett hidratált elaterák
A különnemű spórák elatera szalagjai páratelt térben együtt hidratálódnak, tekerednek és egymáshoz közel pottyannak a nedves környezetbe. Kedvező feltételek esetén egymás közvetlen szomszédjaként indulhatnak fejlődésnek és így könnyebben adódik lehetőség a kifejlődő különböző ivarú előtelepek közötti megtermékenyítésre. A különböző nemű spórák együtt maradása a zavartalan nemzedékváltakozás alapfeltétele.
Újabb vizsgálatok kimutatták, hogy a társaiktól elszakadt magányos spórák fura szalag-pántlika képződményei nem csak a repülést, hanem a földön való haladást is biztosítják. Az elaterák a magányos zsurló spórákon a páratartalomtól függően hidratációjuk és dehidratációjuk révén ugrólábakként is működhetnek. A spórák az elaterák víztartalom változásait követve sajátos bicegő mozgással képesek helyüket változtatni, vagy váratlan, merész mozdulattal a levegőbe ugrani és kedvező szélben tovább vitorlázni a következő nedvesebb páraterű élőhelyre.